Introduction
This paper will attempt to analyze various explanations for the involvement of hominids in the archaeological faunal assemblage from Bed I at Olduvai, specifically the FLK complex, level 22. The analysis will consider evidence regarding the attributes of assemblages created by experimental sequences of access to carcasses by hominids and carnivores versus the actual attributes of the FLK assemblage. The comparison will allow a determination of the sequence of events regarding access to carcasses by hominids through hunting, scavenging, a combination of the two, or variations of the former possibilities.
Experimental assemblages have been created by many researchers in an effort to compare experimental controls to the actual assemblage from FLK 22 in order to determine what sequence of timing access occurred at that site. Results from experiments conducted by Robert J. Blumenschine, Marie M. Selvaggio, and Manuel Dominguez Rodrigo will be examined in this analysis. There seem to be consistent comparative results between the studies that indicates that hominids had secondary access to carcasses after carnivore (inclusive category containing both “hunters” and “scavengers” that may leave similar tooth marks, but differential mark placement) access. Irregularities in the adherence of results to that scenario can be satisfactorily resolved by positing a carnivore-hominid-carnivore three-stage sequence, rather than a hominid-carnivore two-stage sequence in the FLK 22 archaeofauna.
Experimental Controls
Blumenschine
In some of the earliest work involving the construction of experimental faunal assemblages, Robert J. Blumenschine conducted an experiment in 1983 and 1984 in the Serengeti National Park, Tanzania, which involved looking at three separate categories of carnivore and/or hominid involvement (Blumenschine 1988). The experiment consisted of nine carnivore only assemblages, seven “hominid” only assemblages (hammerstone broken assemblages), and twelve hominid-carnivore two-stage sequences, which involved the creation of hammerstone created assemblages that were subsequently allowed to be scavenged (Blumenschine 1988: 488). Blumenschine derived the carnivore only sequences from concurrent studies on the taphonomy of scavenging opportunities (Blumenschine 1986, 1988).
The hominid-carnivore two-stage sequences from experimentally hammerstone broken assemblages, which were defleshed with marrow removed, followed by immediate placement in various settings, and the subsequent recollection of remains after carnivore scavenging occurred (Blumenschine 1988: 485-486). Finally, the “hominid” only assemblage was created by retaining several of the experimentally hammerstone broken assemblages, for comparison, the carnivore scavenged samples, this includes three samples that remained untouched by scavengers due to their setting in riverine woodland sites (Blumenschine 1988: 486, 488).
As should be expected with any early experimental application, this study is full of numerous problems that may or may not have affected results. For example, fragments smaller than two centimetres were restricted from the analysis (Blumenschine 1988: 487), carnivore only were collected from real kill or scavenge sites, and as such, there is no accurate way to determine how much of the actual remains were collected (due to the propensity for hyenas to remove bones from the site) (Blumenschine 1988: 486), and experimental remains were not inclusive of all bones, only long bones and metapodials (Blumenschine 1988: 485-486). However, the gross statistical results are usable, in that they are distinguishable between categories, and are relatively in line with results obtained by other researchers in later experiments.
The results of the experiments are separated into categories that use nomenclature that is also generally adhered to in the other experiments. Carcass taxa are separated into four classes based upon size: size 1 (<50 lbs.), size 2 (50-250 lbs.) size 3 (250-750 lbs.), and size 4 (750-2000 lbs.) (Blumenschine 1986: 642). A fifth category (size 5: >2000 lbs.) is generally not used due to the paucity of available remains for examination, and the rarity in actual collections. Long bone fragments are generally separated into PS (proximal epiphyseal fragments), DS (distal epiphyseal fragments), EPH (epiphyseal fragments that includes PS and DS), NEF (near-epiphyseal fragments), and MSH (midshaft fragments) (Blumenschine 1988: 487; Capaldo 1997: 572). Beyond bone fragment frequencies and cut mark frequencies, Blumenschine’s study is also important in that it shows how bone flakes are caused almost exclusively by hammerstone percussion rather carnivore gnawing (Blumenschine 1988: 494).
Blumenschine’s study quantifies several important features: differences in tooth-marked percentages between carnivore only and hominid-carnivore assemblages, changes in ratios of epiphyseal fragments per whole bone from hammerstone broken stages and carnivore scavenged stages, changes in near-epiphyseal fragments per midshaft fragments between hammerstone broken stages and carnivore scavenged stages, changes in epiphyseal fragments per non-epiphyseal fragments between hammerstone broken stages and carnivore scavenged stages and the association with bone flakes as a result hominid activity rather than carnivore activity in most cases. The results are as follows:
Before carnivore scavenging, hammerstone broken long bones averages 2.2 epiphyseal fragments per original number of bone (both long bones and metapodials). After carnivore scavenging, epiphyseal fragments per original number of bone average 0.06 (Blumenschine 1988: 488).
Table 1
Before After
Original # of
whole bones
epiphyses n epiphyses
bone
epiphyses n epiphyses
bone
Metapodials 22 55 2.5 3 0.14
Non-metapodials 43 86 2.0 1 0.02
Total 65 141 2.2 4 0.06
(Blumenschine 1988: 488) Table 2
The average number of epiphyseal fragments per non-epiphyseal fragment before carnivore scavenging is far greater than after carnivore scavenging, or versus carnivore only assemblages.
Table 2
number of
assemblages epiphyses/
non-epiphyses
S.D.
carnivore only 9 0.03 0.03
hominid only 4* 0.33 0.09
Simulated Sites
open vegetation 10 0.02 0.03
riverine woodland 2 0.41 0.08
fat-depleted 2 0.22 0.09
*not including riverine woodland simulated sites that were not disturbed that are considered hominid only (Blumenschine 1988: 489) Table 3
The average number of near-epiphyseal fragments per midshaft fragment before carnivore scavenging is far less than after carnivore scavenging and far less than the carnivore only assemblages.
Table 3
number of
assemblages epiphyses/
non-epiphyses
S.D.
carnivore only 8 0.29 0.15
hominid only 4 0.07 0.04
Simulated Sites
open vegetation 10 0.18 0.10
riverine woodland 3 0.08 0.07
fat-depleted 2 0.06 0.03
(Blumenschine 1988: 490) Table 4
Carnivore only assemblages have high tooth mark percentages in all bone fragment category, however, in simulated sites where carnivore damage occurred after hominid damage, tooth mark percentages fall in sequentially increasing rates from midshaft to near-epiphyseal to epiphyseal fragments.
Table 4
Size 1 and 2 bovids Size 3 bovids Total
CO SS CO SS CO SS
Total 64/91 44/287 125/140 38/254 189/231 82/541
% 70.3 15.3 89.3 15.0 81.8 15.2
By Limb Segment
epiphyses 2/2 2/3 4/4 1/1 6/6 3/4
% 100 66.7 100 100 100 75.0
near-epiphyseal 7/9 17/34 40/42 23/41 47/51 40/75
% 77.8 50.0 95.2 56.1 92.2 53.3
midshaft 55/80 25/250 81/94 14/212 136/174 39/462
% 68.8 10.0 86.2 6.6 78.2 8.4
adapted from Table 5 (Blumenschine 1988: 491)
The number of bone flakes, as a percentage of the total number of non-epiphyseal bone fragments, is far greater in the hominid only assemblage versus the carnivore only assemblage, showing that bone flakes are a result of hammerstone percussion and only rarely occur during carnivore gnawing.
Table 5
number of
non-epiphyseal
fragments
number of
bone flakes
percentage
hominid only 239 57 23.8
hominid-carnivore 575 109 19.0
carnivore only 229 8 3.6
(Blumenschine 1988: 494) Table 8
Selvaggio
Experiments conducted by Marie M. Selvaggio take a slightly different route. Selvaggio conducted the formation of experimental assemblages, where the carcasses were initially defleshed by carnivore activity, and then was subsequently collected and butchered using stone tools such as could have been used by early hominids (creating a carnivore-hominid assemblage) (Selvaggio 1994: 216-217). The experiments were conducted over a seven-month period during 1989 and 1990 in the Serengeti National Park and Ngorongoro Conservation Area in northern Tanzania (Selvaggio 1994: 217).
Remains were categorized based on the group size of the carnivores involved in the initial consumption of the carcasses: solitary, small group (between two and four animals), and large group (between nine and eleven) (Selvaggio 1994: 218). These remains were then processed for meat scraps and broken up for marrow using lithics commonly found at Plio-Pleistocene sites, such as basalt and quartzite, with some whole limbs left unbroken in order to compare to archaeofaunal assemblages (Selvaggio 1994: 217-218). The resulting bones and bony fragments were separated into subcategories of complete bones (CO), epiphyseal fragments (EPH), near-epiphyseal fragments (NEF), and midshaft fragments (MSH) (Selvaggio 1994: 218).
The separation of data based on group size is an important factor, due to the extrapolation possible of using the data with increasing group size as analogous to what might occur with increasing competition (i.e., changes in assemblages due to the competition may be corrected for when comparing archaeological results to experimental assemblages). The data provided could also be important as a way of testing for hominid scavenging of carnivore kills in a carnivore-hominid two-stage timing sequence, which was not addressed in the initial experimental assemblages by Blumenschine.
Unfortunately, the raw data documented for differences in long bone portions remaining after hominid butchery were unclear and contradictory. Whether this was due to the method of hammerstone breakage to remove marrow (Selvaggio had bones broken from the proximal end first, whereas other researchers broke bones from the diaphyseal portion of the long bones first [Selvaggio 1994: 217; Blumenschine 1988: 485]), or other factors is unknown, but what can be seen is that since the carcasses were not processed after initial abandonment by the primary hunter, the epiphyses were not reduced by bone-crunching scavengers (although some of the carcasses had been hunted by spotted hyenas [Selvaggio 1994: 216]).
What can be gleaned from the data though, is that in assemblages where hominids had secondary access to material before scavengers with the capacity to break down epiphyses for their grease content, there should be large numbers of epiphyseal fragments and few near-epiphyseal fragments (Table 6), due to the breakage caused by hominids in search of marrow (however, how much destruction of these epiphyseal fragments after being discarded by hominids unable to retrieve their grease may make the results rarely comparative to an archaeofaunal assemblage where this kind of timing sequence actually occurred). The most important aspect of the experimental results of Selvaggio is the gross percentages of fragments from portions of the long bones that contained one or more tooth mark, cut mark, or percussion mark.
Thus, useful information gleaned from carnivore-hominid two-stage experimental faunal assemblages include:
In a carnivore-hominid two-stage timing sequence, a high proportion of resulting fragments will be epiphyseal, with a minimum of near-epiphyseal fragments.
Table 6
Long Bone Portions
carnivore
group size number of
carcasses
EPH
%
NEF
%
MSH
%
Total
solitary 15 97 41.28 35 14.89 103 43.83 235
small groups 13 121 44.32 22 8.06 130 47.62 273
large groups 4 9 21.95 11 26.83 21 51.22 41
total 32 227 41.35 68 12.39 254 46.27 549
adapted from Table 1 (Selvaggio 1994: 218)
The percentages of all remains – regardless of carnivore group size – with tooth marks, cut marks, or percussion marks show that: 1) when complete bones remain (which would be more likely than fragmented remains to have flesh scraps) there is a high incidence of hominid cut marks; 2) on fragmented remains recovered by hominids, cut marks will be more likely to occur at the epiphyseal and near-epiphyseal portions (since this is where muscles and tendons attach and will be most likely to survive in some manner); and 3) percussion marks (associated with bone breakage for marrow or spongy trabecular bone) occur relatively evenly over the bones.
Table 7
Long Bone Portions
CO EPH NEF MSH Total
% tooth-marked 49.4 75.0 63.0 47.0 65.0
% cut-marked 67.4 37.0 30.0 17.4 34.1
% percussion-marked N/A 45.0 53.0 35.0 46.0
adapted from Table 2 (Selvaggio 1994: 220)
Dominguez-Rodrigo
The experimental assemblages created by Manuel Dominguez-Rodrigo consist of hominid-carnivore models, carnivore-hominid models, and carnivore-hominid-carnivore models. The experiments were conducted at the Maasai Mara National Reserve and the Galana and Kulalu Ranches near Tsavo East National Park in Kenya (Dominguez-Rodrigo 1997: 672). Long bones and metapodials were again used in this study (as in those previously mentioned above), and were processed by having the flesh removed and marrow processed using lithic materials that would have been available to Plio-Pleistocene hominids (an exception to this is the use of metal knives to remove flesh in two of the experimental assemblages, however, the marks caused by the knives were not counted in the percentages of cut marks in the assemblages) (Dominguez-Rodrigo 1997: 673-674).
Dominguez-Rodrigo used a different nomenclature from Blumenschine and used proximal and distal epiphyses/shaft rather than epiphyseal fragments and near-epiphyseal fragments (Dominguez-Rodrigo 1997: 674). He maintains that this allowed him to distinguish between marks made during disarticulation of skeletal elements versus those made during flesh removal.
However, this does not seem to be explained or used in this study. In any event, the separation of the material into categories for comparison to archaeofaunal assemblages seems haphazard at best, and very uninformative. What can be taken from this study is a comparison between cut mark frequencies on midshaft pieces and combined epiphyseal and near-epiphyseal pieces in different experimental scenarios: hominid only, hominid-carnivore, carnivore-hominid, and carnivore-hominid-carnivore.
In Table 8, the results of Dominguez-Rodrigo are shown. The assemblages are demarcated by H1 (hypothesis 1 – hominid access first), H2 (hypothesis 2 – hominid access second), S1 (stage 1), S2 (stage 2), and S3 (stage 3).
While the correlation of the data presented by Dominguez-Rodrigo are sketchy and seem to have large variance within the sample in each scenario, a tendency for higher cut mark frequency in samples where hominids had first access, decreased when they had secondary access, and minimized even more when carnivores scavenged the remains left behind by hominids can be seen.
Table 8
Cut Mark Frequencies
MSH EPH+NEF Total
H1S1 52.94 61.54 57.53
H1S1/2a 57.14 52.63 55.00
H1S1/2b 70.00 42.86 54.17
H1S1/2c 46.67 37.50 41.94
H1S1/2d 25.00 50.00 29.17
H2S1/2a ** ** 24.39
H2S1/2b 33.33 25.00 28.57
H2S1/2/3a 2.94 12.12 7.46
H2S1/2/3b 14.71 25.00 19.70
**could not distinguish from the data provided
adapted from Table 2 (Dominguez-Rodrigo 1997: 679)
FLK 22 Archaeofauna
The FLK “Zinjanthropus” archaeofaunal assemblage has been at the centre of much of the debate concerning whether early hominids habitually hunted for access to meat, or were primarily scavengers until much later in time. This assemblage comes from the Bed I of Olduvai Gorge, specifically level 22 of the FLK complex, which preserved by far the densest concentration of archaeofauna at the site (Potts 1988: 17-18). The site is particularly well-dated, as it lies between two volcanic tuffs: Tuff IB and Tuff IC. Walter et al. have dated Tuff IB to 1.798 ± 0.004 myr and Tuff IC to 1.761 ± 0.028 myr (Walter et al. 1991). The hominids that have been implicated in the faunal interaction recorded at the site have included A. boisei, A. robustus, H. habilis, and H. erectus (Wolpoff 1999: 365-366), however, the remains of only two of these species has actually been found at the sites of Bed I. These include A. robustus and H. habilis, with the remains of A. boisei specifically having been found within the FLK level 22 material (Potts 1988: 25).
The FLK archaeofauna is important whether or not the mode of hominid interaction of the material was primarily hunting or scavenging since it is evidence that – either way – Plio-Pleistocene hominids were collecting material and moving them to the area of processing. This distinction can be shown by reviewing the nature of the material relative to “natural” accumulations of materials. Natural bone accumulations have several distinguishing features, as described by Potts (1988: 38):
In normal taphonomic accumulation due to attritional mortality, a very low degree of bone concentration results on the landscape. A high concentration in a natural assemblage is 0.62 bones per 100 m2 (Behrensmeyer and Boaz 1980: 74), but the concentration of MNE (minimum number of elements needed to produce the assemblage) at FLK 22 is approximately 1391 bones per 100m2 (Potts 1988: 39-40). Relatively little mixing of bones from different animals occurs, even when predators and scavengers have dispersed bones far away from the carcasses. However, there appears to be an inordinately large MNI (minimum number of individuals) in the relatively small area that level 22 is restricted to in the FLK complex (Potts 1988: 40-41). Also, intrusions of remains from later times is not a consideration due to the volcanic layer capping FLK 22 (some bones from the assemblage even stick up into Tuff IC).
The taxonomic diversity of animals represented in the area tends to be small. At Olduvai, over sixteen taxa of large mammals are found in each faunal assemblage, with woodland species and open plains species occurring in the same assemblages (Potts 1988: 42-43).
Vertebrae and axial limb parts tend to stay in the area near the death site, whereas limb elements tend to be moved (known as the schlepp effect [Bunn and Kroll 1986: 434]). The FLK 22 site is distinguished by a disproportionately small frequency of axial elements relative to limb bones, showing evidence that this was a site where most of the kills did not occur (Potts 1988: 43-44).
In the event of catastrophic (e.g., flood or drought) mortality, there may be large concentrations of bones, but these events leave diagnostic geological evidence. However, the FLK 22 site shows no evidence of any catastrophic mortality event, and the assemblage itself is evidence against this, as almost all remains are fragments, and not articulated pieces (Potts 1988: 44-45).
Each of these factors can be shown to not be the case at FLK 22, and most importantly, while one or more of these factors may not be present in a natural assemblage, the absence of all of them is indicative that artificial collection has taken place. The extensive hominid involvement in the collection (in the form of cut and percussion marks) leads to the conclusion that hominids were responsible for the artificial assemblages.
In order to determine the sequence of events leading to the FLK 22 assemblage by using the experimental data for assemblages in specific scenarios discussed above, the data from FLK 22 must be categorized in a way that allows for comparison with these experimental assemblages. This means that skeletal part profiles for limb bone fragments, cut mark percentages on limb bone fragments, percussion mark percentages on limb bone fragments, and tooth mark percentages on limb bone fragments need to be examined.
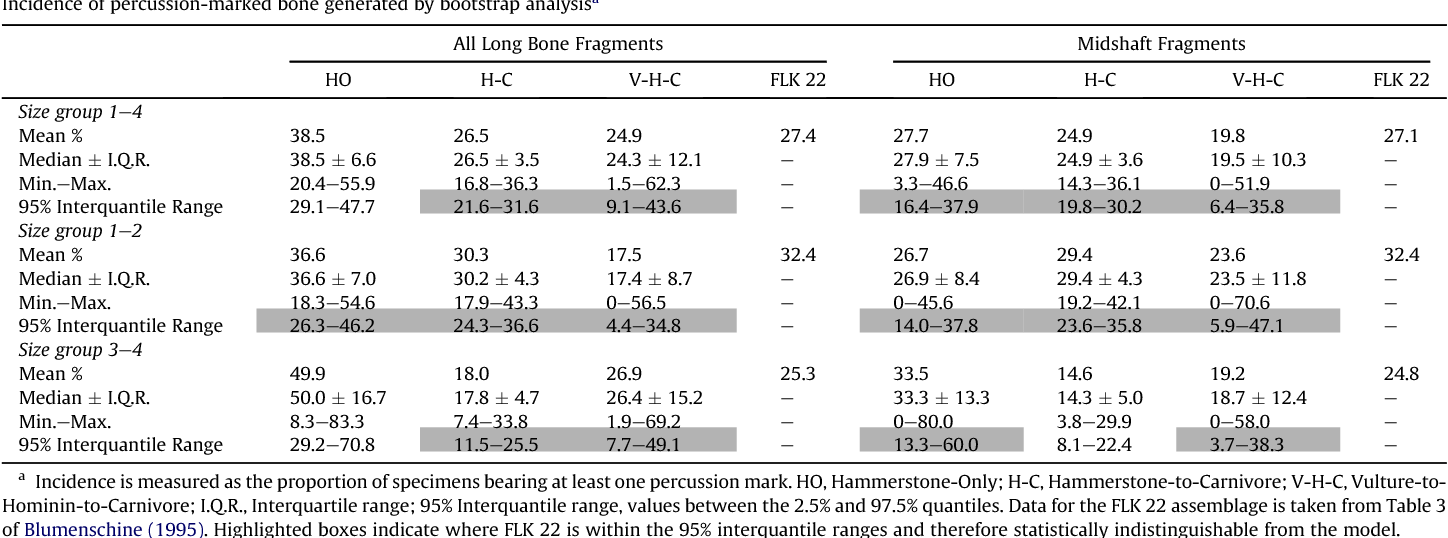
The FLK 22 assemblage consists of approximately 60,000 fragments recovered by Mary Leakey (1971), that have been separated by Bunn and Kroll (1986) into various taxonomic, element, and size categories. While Bunn and Kroll use a larger sample of long bone fragments in their analysis (1467 specimens) (1986: 437), the more detailed study by Blumenschine (1995) of 731 diagnostic long bone fragments will be the sample generally referred to in the remainder of this paper (unless otherwise noted). The comparisons between the experimental assemblages and the FLK assemblage must be taken as a whole, and not necessarily the sum of its parts, and definitely not from any one comparison. Cut mark frequencies, tooth mark frequencies, percussion mark frequencies, and frequencies of long bone portions (EPH, NEF, and MSH).
There is a large variety of data of scenarios in the experimental assemblages including carnivore only assemblages, hominid only assemblages, hominid-carnivore assemblages, carnivore-hominid assemblages, and carnivore-hominid-carnivore assemblages. This data was shown in Table 4, Table 7, and Table 8 above. When compared to the FLK 22 cut mark frequencies, it is obvious that the carnivore only scenario, the hominid only scenario, and the hominid-carnivore scenario do not correlate with the FLK 22 archaeofaunal assemblage. The data concerning carnivore-hominid and carnivore-hominid-carnivore correlation with FLK 22 is less clear, but seem to cluster closer to the carnivore-hominid-carnivore scenario. However, with the small sample size, the differences may be more likely due to sampling error rather than real differences.
Table 9
Cut Mark Frequencies
EPH % NEF % MSH % Total %
FLK 22 37 34.91 28 16.37 72 15.86 137 18.74
adapted from Table 8 (Capaldo 1997: 575)
Tooth mark frequencies measured by Selvaggio (actual frequencies rather than correlation to frequencies shown for FLK 22 by Bunn and Kroll [1986]) for carnivore-hominid experimental assemblages (shown above in Table 7) is almost exactly the same as those seen in FLK 22 (as shown in Table 10 below).
Table 10
Tooth Mark Frequencies
EPH % NEF % MSH % Total %
FLK 22 80 75.47 101 59.06 263 57.93 444 60.74
adapted from Table 3 (Blumenschine 1995: 32-33)
Table 7 above shows the results from a carnivore-hominid experimental assemblage, meaning that all fragments made from the hammerstone destruction of the remains in order to extract the marrow. This means that these numbers for percussion frequencies will be experimental maximums since percussion percentages will drop if carnivores have access to remains after hominids have extracted marrow (as seen in Table 5 above). This may be the result of increased fragmentation in the post-hominid stage increasing the total number of fragments without appreciably increasing the number of percussion scars (or bone flakes which can be used as an indicator of percussion scars), as well as the dispersion of larger fragments, which would be more likely to retain percussion scars (due to retaining the largest percentage of total surface area in the fewest fragments).
Since Table 5 shows bone flakes as a percentage of the total number of non-epiphyseal fragments, one can consider the percentages as a minimum percussion scar percentage for the experimental assemblage – since bone flakes are determined by the presence of a bulb of force and platform on the proximal end of the flake, the total number of bone flakes counted would be unlikely to change much through fragmentation, but the total number of percussion scars could easily be increased through fragmentation (not exponentially, but easily by the degree needed to explain differences in the FLK 22 and experimental assemblages).
Even if fragmentation did not increase the percussion frequency statistically significantly (over that measured by bone flakes), the data seems to clearly indicate (Table 11 below) that the FLK 22 percussion frequencies cluster with a carnivore-hominid-carnivore three-stage timing sequence most closely, followed by correlation with a carnivore-hominid two-stage timing sequence.
Table 11
Percussion Mark Frequencies
EPH % NEF % MSH % Total %
FLK 22 35 33.02 42 24.56 123 27.09 200 27.36
adapted from Table 3 (Blumenschine 1995: 32-33)
Blumenschine’s data (Table 5) shows that: 1) hominid hammerstone activity creates a much higher percussion frequency than carnivore activity, and 2) carnivore activity on a hominid created assemblage will lower the percussion frequency, even though in the processing of the material, the carnivores will actually create some scars. Selvaggio’s data for a carnivore-hominid assemblage can be considered as a hominid-only assemblage since carnivores produce so few scars.
This creates the problem of comparing the percussion percentages in Table 5 for hominid only and in Table 7 for carnivore-hominid (considered hominid only). As stated earlier, Selvaggio used a different method of hammerstone breakage than Blumenschine – Selvaggio started from one end of the shaft and worked down the shaft until all marrow could be removed, while Blumenschine broke initially at midshaft in order to remove the marrow. It seems more likely that hominids would have broken long bones more often from midshaft, although some mixture of methods should be expected, and this can explain why the FLK 22 assemblage has lower percussion frequencies than seen in Selvaggio’s experiments, even lower than can be explained by percussion scar percentage decrease with subsequent carnivore scavenging, and a higher percentage than seen in Blumenschine’s experiments.
The problem with all these data and all determinations from the aforementioned data are that it has not been shown that these percentages are statistically significant or can be considered the norm for these experimental sequences.
That being said, if one considers the mean percentages reported by Blumenschine (Table 5 above) as typical for such assemblages, then one can extrapolate:
H x p + C = HC
Where H is equal to the percussion mark percentage of hominid only assemblages, C is equal the percussion mark percentage of carnivore only assemblages, HC is equal to the percussion mark percentage of hominid-carnivore assemblage, and p is the rate of percussion mark percentage decrease caused by carnivores on a hammerstone created assemblage. Using this equation, one finds that, for Blumenschine’s data, p is equal to approximately 0.65. This means that there is a reduction of 35% in the percussion mark frequency created by hominids when carnivores scavenge the assemblage (although the carnivores themselves add to the final percussion frequency). This type of rate change (if calibrated through extensive experimentation) could be used to explain differences in mark percentages when a different method of breakage is used, resulting in different long bone part proportions and different percussion mark frequencies.
In essence, the gross differences in percussion mark frequencies between experimental assemblages related to archaeofaunal assemblages may be less important than the rate of reduction seen in the experimental assemblages when related to archaeofaunal assemblages and the broad differences between the different experimental scenarios. For example, when one applies the 35% reduction to Selvaggio’s numbers (Table 7 above), the EPH and total percussion mark percentages come very close to FLK 22, with the NEF and MSH greater and lower, respectively. The differences in NEF and MSH frequencies may have to do with the manner of breakage in the experimental assemblage, from one end along the shaft, rather than from midshaft, which I consider to be a more likely place for the greater part of breakage initiation to have occurred.
Thus far, the data given for the experimental scenarios and for FLK 22 seem to show the most likely timing sequence for FLK 22 to be a toss-up between a carnivore-hominid two-stage sequence and a carnivore-hominid-carnivore three-stage sequence. The long bone portion frequencies seem to draw a clear distinction between outcomes of the two scenarios, and squarely place FLK 22 in the three-stage sequence. When the FLK 22 long bone proportions are examined, one finds they do not fit well with either carnivore only, hominid only, carnivore-hominid, or hominid-carnivore assemblages.
Table 12
Long Bone Portions
EPH> % NEF % MSH % Total
FLK 22 106 14.50 171 23.39 454 62.11 731
adapted from Table 3 (Blumenschine 1995: 32-33)
Blumenschine (1988) did not specifically list long bone portion frequencies, but by extrapolating the epiphyseal to non-epiphyseal (NEF and MSH) frequencies from Table 3 (1988: 489) and the near-epiphyseal to midshaft frequencies from Table 4 (1988: 490), one can create long bone portions frequencies for the carnivore only, hominid only, and – using the open vegetation simulated sites (the fat-depleted were a special case scenario, and the riverine woodland were unscavenged, thus were not used in the extrapolation) – hominid-carnivore two-stage sequence assemblages.
These data (Table 13) were created by taking the frequencies for epiphyseal versus non-epiphyseal as meaning EPH/(NEF + MSH), and the near-epiphyseal versus midshaft as meaning NEF/MSH. Second, the MSH in each scenario was assigned the value of 100, allowing the extrapolation of numbers for EPH and NEF, which were subsequently used to form long bone portion frequencies. This allowed accurate (to the degree allowed by the significant figures reported by Blumenschine for the frequencies in his Table 3 and Table 4 [1988]) estimation of long bone portion frequencies regardless of the actual number of bone fragments in each category.
Table 13
Extrapolated Long Bone Portions
*carnivore
hominid hominid-
carnivore
EPH 2.91 24.81 1.96
NEF 21.83 4.92 14.96
MSH 75.26 70.27 83.08
adapted from Table 3 and Table 4 (Blumenschine 1988: 489-490)
*Table 4 listed “8” rather than “9” under the number of assemblages counted in the carnivore sample, this may be a typo (since it wasn’t noted why), but if it is not, it may have slightly affected the numbers, but not appreciably
This shows that hominids create epiphyseal fragments rather than near-epiphyseal fragments (because epiphyses are not broken up very much, and hence do not produce many near-epiphyseal fragments), while carnivores produce few epiphyseal fragments and many near-epiphyseal fragments (because carnivores such as hyenas destroy epiphyses for the grease contained within, creating more near-epiphyseal fragments).
In Blumenschine’s experimental assemblages, when carnivores scavenged the hominid assemblage, there was a massive decrease in epiphyseal fragments and a subsequent increase in near-epiphyseal fragments. This caused an increase in near-epiphyseal frequency due to the increase near-epiphyseal fragments, an increase in midshaft frequency due to the total decrease in fragments relative to the number of midshaft fragments, and a decrease in the epiphyseal frequency due to the loss of epiphyseal fragments.
When you compare the carnivore-hominid assemblage (Table 6 above) to the hominid only assemblage (Table 13 above) – considered as pre-carnivore scavenged assemblages – one can see that in a carnivore-hominid-carnivore assemblage, the resulting long bone frequencies should be higher than the hominid-carnivore in the EPH and NEF categories, and lower in the MSH category. This is due to the higher initial EPH frequency causing a higher resulting EPH frequency [24.81(H) ==> C = 1.96(H-C), thus, 41.3(C-H) ==> C > 1.96(C-H-C)], the higher initial NEF frequency causing a higher resulting NEF frequency [4.92(H) ==> C = 14.96(C-H), thus, 12.4(C-H) ==> C > 14.96(C-H-C)], and a lower initial MSH frequency causing a lower resulting MSH frequency [70.27(H) ==> C = 83.08(H-C), thus, 46.3(C-H) ==> C < 83.08(C-H-C)]. This is seen in the FLK 22 assemblage, indicating a carnivore-hominid-carnivore three-stage timing sequence as the method of processing.
Conclusions
It has been shown in the discussion above, that the archaeofaunal assemblage is definitely a consequence of hominid interaction through transportation of the material from outside sources, and not a natural accumulation (contra Binford 1981). The archaeofaunal assemblage from FLK 22 itself has obviously been utilized by hominids as shown by the extensive cut marks and percussion marks. The makeup of the assemblage with regards to long bone portion frequency (EPH, NEF, and MSH) clearly indicates that the assemblage was created by the interaction of carnivore only material and material resulting from carnivore and hominid activity. The bone portion frequencies seem to clearly show that after hominid interaction with the material, there was scavenging of the remains by carnivores. This leads to the conclusion that the FLK 22 Zinjanthropus archaeofaunal assemblage is the result of a carnivore-hominid-carnivore three-stage timing sequence.
A question can now be made as to how much meat was on the remains scavenged by the hominids in question. One can accept the argument made by Binford (1988) and Binford et al. (1988) that the hominids were very marginal scavengers with access to only marrow and inconsequential amounts of meat scraps, that of Blumenschine (1995) that the hominids had access to very little residual meat (i.e., were not removing large amounts of flesh, but more than indicated in Binford’s model), or that hominids had access to large amounts of flesh, as suggested by Oliver (1994), Cavallo and Blumenschine 1989, or Capaldo (1997).
Regardless, it is highly unlikely using the experimental assemblages created by Blumenschine, Selvaggio, and Dominguez-Rodrigo as a baseline, that hominids had first access to the material. Looking at the cut mark frequencies on the FLK 22 assemblage where there are no associated tooth marks on the fragments (Table 14 below) one can also see this, as the percentages are extremely low. When hominids have first access to a carcass, carnivores do not bother with non-epiphyseal fragments, as they hold no nutritional value with the meat and marrow removed (Blumenschine 1988: Table 6). Thus, the low frequency of cut marked only midshaft fragments indicates that hominids rarely, if ever, had first access to carcasses or extremely early access to carcasses.
Table 14
Cut Mark Only Frequencies
EPH % NEF % MSH % Total %
FLK 22 3 2.83 12 7.02 20 4.41 35 4.79
adapted from Table 8 (Capaldo 1997: 575)
Therefore, it seems likely that the FLK 22 archaeofaunal assemblage was created by hominids scavenging kills after carnivore abandonment – rather than confrontational scavenging – since confrontational scavenging would likely lead to earlier access to meaty limbs, and hence more cut marks with no tooth marks on them. This sequence was completed with carnivores scavenging the remains, particularly the epiphyseal fragments for their bone grease.
Bibliography
Behrensmeyer, A. K., and D. Dechant Boaz. 1980. “The recent bones of Amboseli Park, Kenya, in relation to East African paleoecology.” In Fossils in the Making, ed. by A. K. Behrensmeyer and A. Hill, pp. 72-92. Chicago: University of Chicago Press.
Binford, Lewis R. 1981. Bones: Ancient Men and Modern Myth. San Francisco: Academic Press.
Binford, Lewis R. 1988. “Fact and Fiction about the Zinjanthropus Floor: Data, Arguments, and Interpretations.” In Current Anthropology, vol. 29, no. 1, pp. 123-149.
Binford, Lewis R., M. G. L. Mills, and Nancy M. Stone. 1988. “Hyena Scavenging Behavior and Its Implications for the Interpretation of Faunal Assemblages from FLK 22 (the Zinj Floor) at Olduvai Gorge.” In Journal of Anthropological Archaeology, vol. 7, pp. 99-135.
Blumenschine, Robert J. 1986. “Carcass Consumption Sequences and the Archaeological Distinction of Hunting and Scavenging.” In Journal of Human Evolution, vol. 15, pp. 639-659.
Blumenschine, Robert J. 1988. “An Experimental Model of the Timing of Hominid and Carnivore Influence on Archaeological Bone Assemblages.” In Journal of Archaeological Science, vol. 15, pp. 483-502.
Blumenschine, Robert J. 1995. “Percussion marks, tooth marks, and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania.” In Journal of Human Evolution, vol. 29, pp. 21-51.
Bunn, Henry T., and Ellen M. Kroll. 1986. “Systematic butchery by Plio/Pleistocene hominids at Olduvai Gorge, Tanzania.” In Current Anthropology, vol. 27, no. 5, pp. 431-452).
Capaldo, Salvatore D. 1997. “Experimental determinations of carcass processing by Plio-Pleistocene hominids and carnivores at FLK 22 (Zinjanthropus), Olduvai Gorge, Tanzania.” In Journal of Human Evolution, vol. 33, pp. 555-597.
Cavallo, John A., and Robert J. Blumenschine. 1989. “Tree-stored leopard kills: expanding the hominid scavenging niche.” In Journal of Human Evolution, vol. 18, pp. 393-399.
Dominguez-Rodrigo, Manuel. 1997. “Meat-eating by early hominids at the FLK 22 Zinjanthropus site, Olduvai Gorge (Tanzania): an experimental approach using cut-mark data.” In Journal of Human Evolution, vol. 33, pp. 669-690.
Leakey, Mary D. 1971. Olduvai Gorge, vol. 3, Excavations at Beds I and II. 1960-1963. Cambridge: University of Cambridge Press.
Oliver, J. S. 1994. “Estimates of hominid and carnivore involvement in the FLK Zinjanthropus fossil assemblage: some socio-ecological implications.” In Journal of Human Evolution, vol. 27, pp. 267-294.
Potts, Richard. 1988. Early Hominid Activities at Olduvai. New York: Aldine De Gruyter.
Selvaggio, Marie M. 1994. “Carnivore tooth marks and stone tool butchery marks on scavenged bones: archaeological implications.” In Journal of Human Evolution, vol. 27, pp. 215-228.
Walter, R. C., P. C. Manega, R. L. Hay, R. E. Drake, and G. H. Curtis. 1991. “Laser-fusion 40Ar/39Ar dating of Bed I, Olduvai Gorge, Tanzania.” In Nature, vol. 354, pp. 145-149.
Wolpoff, Milford. 1999. Paleoanthropology. Boston: McGraw-Hill.